Biosonar, diving and movements of two tagged white-beaked dolphin in Icelandic waters
In courtesy of Dr. Marianne H. Rasmussen
Abstract
For the first time bio-logging tags were attached to free-ranging white-beaked dolphins, Lagenorhynchus albirostris. A satellite tag was attached to one animal while an acoustic A-tag, a time-depth recorder and a VHF transmitter complex was attached to a second dolphin with a suction cup. The satellite tag transmitted for 201 days, during which time the dolphin stayed in the coastal waters of western Iceland. The acoustic tag complex was on the second animal for 13 hours and 40 minutes and provided the first insight into the echolocation behaviour of a free-ranging white- beaked dolphin. The tag registered 162 dives. The dolphin dove to a maximum depth of 45 m, which is about the depth of the bay in which the dolphin was swimming. Two basic types of dives were identified; U-shaped and V-shaped dives. The dolphin used more time in U-shaped dives, more clicks and sonar signals with shorter click intervals compared to those it used in V-shaped dives. The dolphin was in acoustic contact with other dolphins about five hours after it was released and stayed with these for the rest of the tagging time. Possible foraging attempts were found based on the reduction of click intervals from about 100 ms to 2-3 ms, which suggests a prey capture attempt. We found 19 punitive prey capture attempts and of these 53 % occurred at the maximum dive depth. This suggests that more than half of the possible prey capture events occurred at or near the sea bed.
Keywords: Bioacoustics, Echolocation, Marine mammals, White-beaked dolphins, Lagenorhynchus albirostris

A schematic drawing of the acoustic A-tag with the two hydrophones the VHF transmitter and suction cup (above) and a photo of the actual tag with the antenna of the VHF visible and the time depth recorder (DST-milli, Star-Oddi, Iceland, not visible) used on the female dolphin.
1. Introduction
White-beaked dolphins (Lagenorhynchus albirostris) are only found in the North Atlantic (Reeves et al., 1999). They are the most common delphinid species in Icelandic waters (Gunnlaugsson et al., 1988; Gunnlaugsson and Sigurjónsson, 1990; Pike et al., 2009; Rasmussen and Miller, 2002; Víkingsson and Ólafsdóttir 2004). The acoustic repertoire of white-beaked dolphins consists of whistles (Rasmussen and Miller, 2002; Rasmussen et al, 2006) and clicks (Rasmussen and Miller, 2002; Rasmussen et al, 2002). North Atlantic Sigthings Surveys have been conducted in Icelandic waters since 1986, usually in July. The distribution of dolphins seemed consistent from aerial surveys conducted in 1986, 1987, 1995 and 2001 with dolphin sightings concentrated in the south-western, north-eastern and south-eastern parts of Iceland, in relatively coastal waters (Pike et al., 2009). Magnusdottir (2007) studied the annual distribution of white-beaked dolphins around Reykjanes Peninsula and she found white-beaked dolphins in Faxaflói Bay also during the winter. The corrected abundance estimate was 31,653 (95 % CI 17,679 – 56,672) from the 2001 survey (Pike et al., 2009). Whale watching started in Iceland in 1991 (O’Connor et al., 2009) and white-beaked dolphins are among the most commonly sighted species on whale watching tours in Iceland (Rasmussen 1999; Salo, 2004; Magnusdóttir, 2007; Bertulli, 2010).

A satellite tagtag (SPOT5, Wildlife Computers, WA, USA) was attached to the dorsal fin to the male dolphin.
Rather little is known about the movements of white-beaked dolphins in Icelandic waters. Photo-identification studies have been conducted in Faxaflói Bay from 1997 – 2010 (Rasmussen, 1999; 2004, Magnusdottir, 2007; Bertulli, 2010). Rasmussen and Jacobsen (2003) showed that 12-20 % of the dolphins had markings that could be used for individual identification and the same individual was re-sighted up to nine times during a season. Bertulli (2010) found a total of 28 re-sightings of the same individual in Faxaflói Bay between 2007 and 2009. So far only a few re-matches of the same individual between areas have been found. One dolphin was photographed in Breiðafjörður and re-photographed in Skjálfandi Bay, Northeast Iceland. The dolphins moved about 300 km in just a few days (Tetley et al., 2006). Three other individuals were photographed in Faxaflói Bay and re-sighted in Skjálfandi Bay in 2009 (Bertulli, 2010).
Satellite tags and radio tags have been deployed on many species of small cetaceans. Perrin et al. (1979) deployed various types of radio tags on spotted dolphins (Stenella attenuata) and spinner dolphins (Stenella longirostris). Scott et al (1990) reviewed various methods for the tagging of small cetaceans and described the tagging of bottlenose dolphins (Tursiops truncates), Hawaiian spinner dolphins, common dolphins (Delphinus delphis), and white-sided dolphins (Lagenorhynchus acutus). Later Mate et al., 1994 and Mate et al., 1995 deployed satellite tags on a white-sided dolphin and a bottlenose dolphin. The total estimated straight-line distance travelled by the white-sided dolphin was 308.9 km (Mate et al., 1994) and the overall distance travelled by bottlenose dolphins was at least 581 km (Mate et al., 1995). Satellite tags have been deployed on other species of odontocetes like on harbour porpoises (Phocoena phocoena) (Read and Westgate, 1997), belugas (Delphinapterus leucas) (Suydam et al., 2001; Hobbs et al., 2005) and narwhals (Monodon monoceros), (Dietz and Heide-Jørgensen, 1995; Dietz et al., 2001).
The diving behaviour of white-beaked dolphins is unknown. However, various time depth recorders (TDRs) have been used to study diving in other odontocetes (Baird et al., 2002; Baird et al., 2006; Baird et al., 2008; Otani et al., 1998; Westgate et al., 1995; Teilmann et al., 2007; Johnson et al., 2009). Hooker and Baird (2001) published a review on the subject describing the diving behaviour of 13 species of odontocetes. Of these, four were dolphin species: Common dolphins (Delphinus delphis), Heaviside’s Dolphin, (Cephalorhynchus heavisidii), Pantropical spotted dolphins (Stenella attenuate) and Atlantic spotted dolphins (Stenella frontalis).
Otani et al. (1998) and Westgate et al. (1995) described different types of dives for harbour porpoises. Otani et al. (1998) report V-shaped dives without a defined bottom time, which they classified as transit and travelling dives. Westgate et al. (1995) described U-shaped dives with a flat bottom phase lasting seconds to several minutes, which are believed to be foraging dives. Scott and Chivers (2009) recorded night-time U-shaped dives for spotted dolphins with rapid changes in depth (“wiggles”) while at the bottom of the dive and suggested these occurred during the pursuit of prey.
Sigurjónsson and Vikingsson (1997) estimated that white-beaked dolphins in Icelandic waters feed mainly on fish (95 %) and squid (5 %). From indirect data Sæmundsson (1939) concluded that capelin and herring were the most common prey species in Icelandic waters. While confirming these two species as a part of the diet of white beaked dolphins, more recent studies identified larger fish such as haddock (Melanogrammus aeglefinus), cod (Gadus morhua), and saithe (Pollachius virens) as the most important constituents in the white-beaked dolphins’ diet (Víkingsson and Ólafsdóttir 2004). Inside Faxaflói Bay they also feed on sandeels (Ammodytidae sp.) (Rasmussen, 1999; Rasmussen and Miller 2002; Rasmussen, 2004; Bertulli, 2010).
Presumably they use their echolocation clicks to find prey and to navigate. Some properties of these signals have been investigated with a hydrophone or hydrophone arrays operated from small boats (Rasmussen and Miller, 2002; Rasmussen et al., 2002; and Rasmussen et al., 2004). Attaching acoustic tags on free- ranging dolphins is an effective method for studying how they use echolocation during prey capture. The most commonly used acoustic tags on small cetaceans, like porpoises, are A-tags, developed in cooperation with the National Research Institute of Fisheries Engineering in Japan (Akamatsu et al., 2005c). D-tags, developed at Wood Hole Oceanographic Institution in USA (Johnson and Tyack, 2003), have beendeployed on larger odontocetes among other cetaceans. The A-tag has two hydrophones and necessary electronics to function as a 2-channel, high frequency, event-recorder to capture the ultrasonic clicks of odontocetes. A-tags have been used to describe echolocation behaviour of wild harbour porpoises (Akamatsu et al. 2007; Linnenschmidt et al., in press) and finless porpoises (Neophocaena phocaenoides) (Akamatsu et al., 2000, 2005a, 2005b, 2010). The D-tag samples the signal waveform at a high rate on two channels, among other behavioural parameters depending on the aim of the project. D-tags have been used to record both communication sound for example in narwhals (Shapiro, 2006), and to study foraging and echolocation behaviour for example in sperm whales (Physeter macrocephalus) (Miller et al., 2004), beaked whales (Zimmer et al., 2005, Johnson et al., 2004, Johnson et al., 2006), short-finned pilot whales (Globicephala macrorhynchus) (Aguilar Soto et al., 2008), and captive harbor porpoises (DeRuiter et al., 2009).
Often when interpreting the results from A-tags and D-tags it has been assumed that the echolocation behaviour of foraging toothed whales can be divided into search (long click intervals), approach (decreasing click intervals) and terminal or catch phases (very short intervals) often called ‘buzzes’. These three phases were first described for echolocating bats when capturing insect prey (Griffin, 1958), but can also be applied to foraging odontocetes like captive harbour porpoises when capturing fish (DeReuiter et al., 2009; Miller, 2010; Verfuss et al., 2009). Recordings of un-instrumented free-ranging narwhals also reveal click sequences that suggest foraging (Miller et al., 1995). Most of the odontocetes equipped with acoustic tags show click phases indicative of prey capture (Johnson et al. 2004, 2007; Linnenschmidt et al., in press; Miller et al., 2004; Madsen et al., 2005).
Many studies on echolocation behaviour of captive odontocetes especially the bottlenose dolphins are summarized in Au (1993) and Au (2000). There are also studies describing the echolocation of wild dolphins using hydrophone-arrays (Au and Herzing, 2003; Rasmussen et al., 2002; Wahlberg et al, 2011). But, no studies exist describing the echolocation behaviour of free-ranging dolphins using acoustic tags.
Consequently, one aim of this study was to describe the acoustic and dive behaviour of a free-ranging dolphin using an attached A-tag and dive recorder. A second aim was to record the movements of a second white-beaked dolphin. This animal wore a satellite tag to record seasonal movements in Icelandic waters. The dolphins were captured in a hoop net with permission from Icelandic authorities (Nachtigall et al. 2008).
2. Materials and Methods
The project was conducted in July and August 2006 in Faxaflói Bay, Southwest Iceland (see Fig. 3) for the purpose of capturing wild white-beaked dolphins to study hearing, movements and acoustic behaviour (Nachtigall et al., 2008; Mooney et al., 2009). We modified a fishing vessel with a platform on the bow for catching dolphins and a holding tank for maintaining dolphins during hearing studies and tagging. We spent around 340 hours on the water and captured two dolphins using a hoop net. The dolphins were placed in a stretcher and lifted on-board into a tank with dimensions of 1 x 1 x 3.7 m for the hearing experiment. Sounds were projected in front of the dolphin and suction cup electrodes were places on the head of the animal as well as a reference electrode on the dorsal fin. Before release the dolphins were equipped with tags (Nachtigall et al. 2008).
2.1. The acoustic tag package
The acoustic tag we used (A-tag, W20-AS, 2-channel, drift: 1 s per day, Little Leonardo, Tokyo, Japan) functions as an ultrasonic event recorder that registers the sound pressure (peak to peak (p-p) re 1 μPa) and the exact time of detection at each of two hydrophones, that are spaced about 120 mm apart. Signals are band pass filtered (55 kHz to 235 kHz) and a hardware detection threshold is set at 142 dB (p-p re 1 μPa). Only sound pulses exceeding this intensity at the front hydrophone trigger events that are stored. If the second hydrophone is also triggered by the same sound pulse, the time arrival difference between the hydrophones is stored with 271 ns resolution. If the second hydrophone is not triggered the time difference is set to zero. Sound pressure and time arrival differences are stored every 0.5 ms. White-beaked dolphins emit clicks with source levels between 189 to 219 dB μPa (Rasmussen et al. 2002) A received level of 189 dB is 47 dB above the trigger level of the A-tag (142 dB), corresponding to other dolphins clicking at a distance of about 100 m (at 100 kHz). For a source level of 219 dB the distance would be 500 m, which is 77 dB above trigger level of the A-tag.
The total recording time is battery limited to approximately 40 h. All components fit into a cylindrical waterproof aluminium housing measuring 21 mm in diameter x 122 mm in length weighing 77 g. The A-tag has been tested successfully to a water depth of 200 m. Detailed information on the A-tag is available in Akamatsu et al., 2005c and Akamatsu et al., 2007.
We imbedded the A-tag in a float (expanded polyvinyl chloride, Klegecell #55, pressure resistant to 80 N/cm2, Kaneka, Japan) for positive buoyancy after the detachment. Also embedded into the float was a time and dive depth recorder (TDR) (DST-milli, Star-Oddi, Iceland). Resolution and accuracy of the depth recorder was 0.03 % of full scale (900 m, i.e. +/-27cm) and +/-0.4% of depth reading, respectively.
We also attached a VHF transmitter (MM130, Advanced Telemetry Systems, MN, USA) for locating the float after detachment. The total weight of tag with dive depth recorder and VHF transmitter was 251 g.
2.2. Animal 1
A 209 cm female white-beaked dolphin was caught on 2 August 2006 (Table 1). After testing its hearing (Nachtigall et al., 2008), it was equipped with the acoustic tag package using a suction cup (see Fig. 1). The animal was released outside Garður harbor (64 ̊N, 22 ̊W, the green dot in Fig. 3) at 18:00 on 2 August 2006 very close to where it was caught. The 28 m high lighthouse at Garðskagi was used for tracking radio signals. At 08:00 am the following day continuous VHF signals were received indicating that the tag had detached and floated on the sea surface. The tag was recovered about 6 NM northeast of the release site (Fig. 3 the star). The suction cup tag was attached on the animal for 13 hours and 40 minutes.
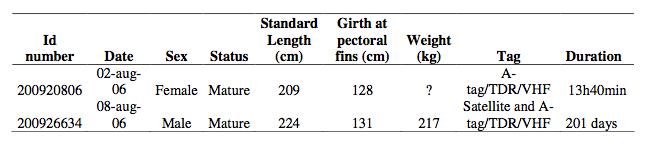
Basic data on the two white-beaked dolphins captured and tagged in this study.
2.3. Animal 2
A second white-beaked dolphin, a 224 cm male, was caught on 8 August 2006 (Table 1). A full audiogram determined by measuring auditory evoked potentials was made on animal 2 (Nachtigall et al., 2008). A satellite transmitter (SPOT5, Wildlife Computers, WA, USA) with an oval shape (measuring 10 cm x 3.5 cm x 1 cm, length x wide x height) was attached to the dorsal fin using two 5 mm silicone covered delrin pins and iron nuts. A picture of the dorsal fin with the satellite tag is shown in Fig. 2. A description of the general attachement methods can be found in Teilmann et al (2007) and Svegaard et al (2011).
The satellite transmitter was set to transmit every 45 s when at the surface. The pre-set maximum of 250 transmissions per day (equals 4-7 hours) applied for the month of August and then 250 transmissions every other day for the remaining time. Satellite positions were obtained from the ARGOS satellite system and had an accuracy of less than a hundred meters to a few kilometres. The satellite tag was designed to remain on the animal for about a year until the iron nuts corroded and the pins fell out. We received positions of animal 2 for 201 days (Fig. 3).
2.4 Dive and Acoustic Data Analysis
The TDR data was synchronized with the A-tag data using the pattern of surface splash noise and the internal clocks with Igor Pro spread sheet software (Wave Metrics, USA). A dive was defined as the period between two surfacings when the depth exceeded 2 m. Dives less than 1 s were not analyzed. “Bottom-time” was defined as in Hooker and Baird (2001) as the time the dolphin spent in a zone between the maximum dive depth and 85% of maximum depth. The dives were divided into two categories: U-shaped dives and V-shaped dives. A U-shape dive was defined as a dive where the time spent at the bottom of the dive, termed the “bottom- time”, was more than 50 % of the total dive time. We defined a V-shaped dive as having a “bottom-time” less than 50 % of the total dive time. The term “bottom-time” refers to the time spent at the bottom of the dive and not the time spent at the sea bottom. The inner part of Faxaflói Bay is quite shallow with water depths between 40 and 50 m.
The A-tag only records events with ultrasonic frequencies and biosonar signals were extracted by reducing transient noise components according to the following procedure. Clicks having a received level of less than 142 dB re 1 μPa peak-to-peak were eliminated. Click trains containing fewer than 5 clicks were also eliminated as echolocating dolphins usually produce longer sequences of ultrasonic pulses (Au 1993). If the click interval being measured was not between one-third and three times that of the previous click interval this click was considered noise and eliminated. Inter pulse intervals in a click train changed relatively smoothly compared with randomly occurring noise such as cavitations. The criteria used in these analyses were chosen conservatively because such a quick change of inter pulse interval rarely happens in biosonar pulse trains. In addition, this filtering allows taking into account a series of pulses and even a missing pulse in a train, which could occur when the received level is close to the detection threshold level.
The two hydrophones allowed for measuring time of arrival differences and calculating a bearing angle to a neighbouring dolphin vocalizing in the vicinity of the tagged animal. The relative angle between the tag and another conspecific (or other odontocete) can vary slightly owing to small changes in time differences caused by head movements of the tagged animal (Akamatsu et al., 2010).
Statistics were done in Microsoft Excel ver. 14.2.2. Differences were considered significant when the probability (p) was equal to or less than 0.05.
3. Results
3.1. Movements of animal 2
We captured a male dolphin in Faxaflói Bay on 8 August 2006 and attached a satellite transmitter to its dorsal fin. Positions of the dolphin were registered until 24 February 2007, seven months later, and in total 201 days. The areas visited by the dolphin were arbitrarily divided into four parts: Westfjords, Breidafjörður, Faxaflói and the South Coast (Fig. 3). The dolphin always stayed within the 200 m depth contour. It travelled a minimum total distance of 5,280 km with a mean speed of 4 km/hr. The longest monthly distance travelled was in August (1,090 km), where it swam from Faxaflói to the Westfjords and shortest was in September 2006 (559 km), where it stayed around the Westfjords (Table 2). The mean swimming speed for each month ranged from 3.5 km/hr to 5 km/hr.
After the dolphin was tagged in August it stayed in Faxaflói six days before moving north to Breiðafjörður and the Westfjords. The dolphin moved south again to Breiðafjörður and Faxaflói in October. It stayed along the south coast in December before swimming back to Faxaflói. From Faxaflói the dolphin had another trip north passing Breiðafjörður to the Westfjords and south again to Faxaflói in January. The dolphin went along the south coast and was close to the Vestmannaeyjar Islands in February 2007 before going back to Faxaflói, where the tag stopped sending (Fig. 3). In total the dolphin travelled the longest distance and spent most of its time in Faxaflói (44 % of the time). It spent 28 % of the time around the Westfjords, 18 % along the south coast and 10% in Breiðafjörður.
3.2. Diving behavior of animal 1
We captured a female dolphin also in Faxaflói Bay and attached an A-tag, a TDR and a VHF transmitter packaged to a suction cup (Fig. 1). The tag package remained on the animal for 13 h and 40 m during which time it made 142 dives. It spent 18 % of the time close to the surface (0-2 meter) and 82 % of the time diving to depths greater than 2 m (Fig. 4). The dive duration varied between 6 s and 128 s and the bottom time varied between 2 s and 78 s (mean 28 s ± 21 s sd). The dolphin was mainly making shallow dives during the first hour (down to 8 meter), but after 55 minutes the dolphin dove down to 34 m. The maximum dive depth was 45 m, which is about the depth of the bay. The highest rate of diving occurred between 2 am and 7 am with a maximum of 24 dives per hour around 4 am. Diving was less frequent from 7 pm to 1 am.
The dolphin made two types of dives: U-shaped and V-shaped and examples are shown in Fig. 5 and summaries in Table 3. There were not significantly more U- shaped dives than V-shaped dives and the time spent in the two types of dives was likewise not significant (Table 3). The mean time the dolphin spent at the bottom of the dive was significantly longer for U-shaped dives while the time used in descent and ascent was the same for the two types of dives.
3.3. Acoustic behavior of animal 1
The A-tag registered acoustic events for the entire 13 h and 40 min the tag was attached to the dolphin. The acoustic data on the tag revealed how the animal used sonar during diving, presumed foraging, and when the animal was in the company of other dolphins. The difference in arrival time at the two hydrophones can be used to determine the bearing to a neighbouring animal. Acoustic contact with another dolphin was first recorded after 4.5 hours following release, and during the rest of the time the tag was on the dolphin. Thus, our tagged dolphin was in acoustic contact with other dolphins most of the time (Fig. 4). Figure 6 shows an example where a dolphin was swimming around our tagged dolphin. The dotted lines on the upper trace show another dolphin passing our tagged dolphin, where plus means the sound arrived at the front hydrophone first and minus means sound arrived at the back hydrophone first. As the neighbouring dolphin approached the tagged dolphin the inter pulse intervals decreased to 2-3 ms indicating a close approach.
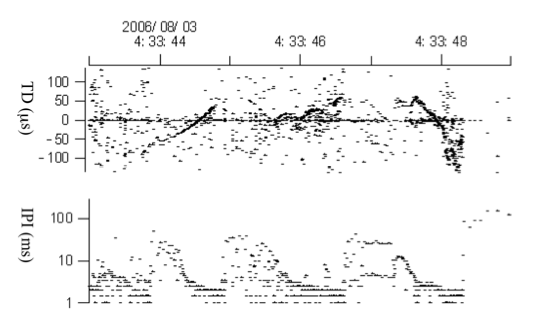
Examples of acoustic contact between the tagged animal and another animal. The other individual(s) swam around the tagged animal. Note that the interpulse (click)- interval (IPI) is very short (2-3 ms) at the end of the click sequence. This suggests the other animal approached the tagged animal to within a short distance. TD is time difference between the two A-tag hydrophones where plus means the sound arrived at the front hydrophone first (see methods).
The dolphin used sonar signals with approximately 200 ms pulse intervals at the surface. In contrast, the pulse intervals during diving were approximately 100 ms or about half that produced at the surface. The dolphin used significantly fewer clicks in V-shaped dives but at a higher rate in general compared to clicks used in the U- shaped dives (Table 3). There was no significant difference in mean click rate or the click rate used during the bottom time and during the ascent for the two types of dives. The dolphin used significantly longer click intervals at the bottom of V-shaped dives (Table 3).
We used the occurrence of shortening click intervals when looking for possible foraging attempts. Click intervals that decreased from about 100 ms to 2-3 ms could indicate a foraging attempt. We show an example of a possible foraging event in Fig. 7, which happened at a depth of 18 m. The nineteen possible foraging events were randomly distributed between V-shaped (10) and U-shaped dives (9). The mean maximum dive depth for presumptive foraging was 25 m in V-shaped dives and 28 m in U-shaped dives. Fifty three percent (10 of 19) of the presumed foraging events happened at the bottom of the dive at a depth of 5 to 37 m and the rest occurred on either the descent or ascent (Fig. 8). There was significantly more presumptive prey capture activity during the descent/ascent in V-shaped dives compared to U- shaped dives (p = 0.032). There was no difference in probable prey capture activity near the bottom of V-shaped or U-shaped dives (p = 0.074). All presumptive foraging occurred the morning between about 1:30 am and 7 am (Fig. 4), when the ambient light was low.
4. Discussion
4.1 Movements of a satellite tagged white-beaked dolphin
This is the first time a satellite tag has been placed on a white-beaked dolphin. The tag transmitted for 201 days. This is in contrast to satellite tagging of Atlantic white-sided dolphins (Lagenorhynchus acutus) in the Golf of Maine, where the tag transmitted for six days (Mate et al., 1994) and for bottlenose dolphins (Tursiops truncatus) in Florida where transmission was 25 days (Mate el al, 1995). The satellite transmitters on Atlantic spotted dolphins (Stenella frontalis) in the Gulf of Mexico transmitted for 24 days (Davis et al., 1996). Our white-beaked male dolphin travelled a mean minimum distance of 26.3 km/day (Table 2), which is much shorter than mean distance of between 111-248 km/day by pelagic spotted dolphins (Stenella attenuata, Scott and Chivers, 2009). The mean swimming speed for our white-beaked dolphin per month ranged from 3.5 km/h to 5 km/h, but these are only rough measurements. For comparison 17 tagged spotted dolphins had individual speeds of between 5.2 km/h and 10.4 km/h (Scott and Chivers, 2009).
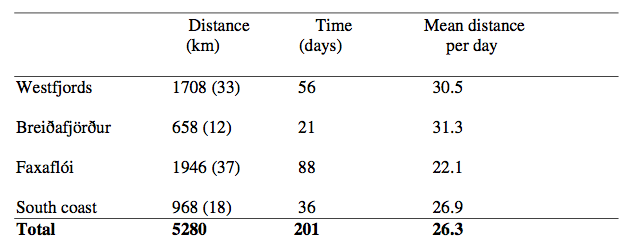
Distance travelled and time spent in the four different areas for the male white-beaked dolphin. Numbers in parentheses show the percent of total distance spend in each area. The animal was tagged for 201 days. (See Figure 3 for a definition of the areas.)
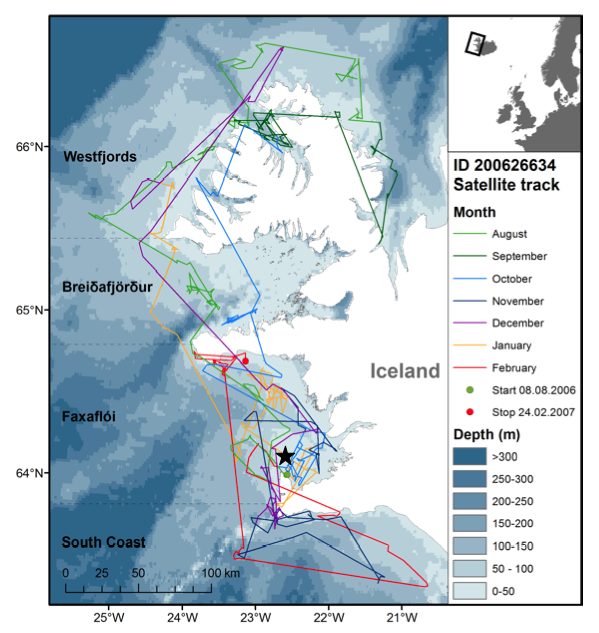
A small map of Iceland with an enlargement of western Iceland showing the movements of a white-beaked dolphin equipped with a satellite transmitter. Transmissions started on 8 August 2006 (green dot, Garður harbor) and stopped on 24 February 2007 (red dot). Different color lines show the distance covered by the tagged animal in different months. The area west and south of Iceland was divided in four parts corresponding to the location: Area 1: The Westfjords, Area 2: Breiðafjörður, Area 3: Faxaflói and Area 4: South Coast. An acoustic A-tag was placed on a second dolphin at the green dot. The acoustic tag was recovered approximately 6 NM northeast of the lighthouse in Garður (the star). (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
White-beaked dolphins are typically found over continental shelves in the North Atlantic. Our satellite tagged white-beaked dolphin stayed within the 200 meter depth contour or closer to shore (Fig. 3). This is in agreement with sightings of white- beaked dolphins from whale watching vessels around Iceland from April – October (Rasmussen, 1999; Rasmussen, 2004; Salo, 2004; Bertulli, 2010) and also sightings of coastal white-beaked dolphins during summer in the North Atlantic Sightings Surveys (Pike et al., 2009; Gunnlaugsson et al., 1988; Gunnlaugsson and Sigurjónsson, 1990). Magnusdóttir (2007) observed white-beaked dolphins in winter in Faxaflói and along the South Coast of Iceland and white-beaked dolphins have also been sighted in Faxaflói during winter whale watching in Faxaflói (www.elding.is). By-catch and stranding records also confirm the year-round presence of white-beaked dolphins (The Icelandic Marine Research Institute, unpublished data).
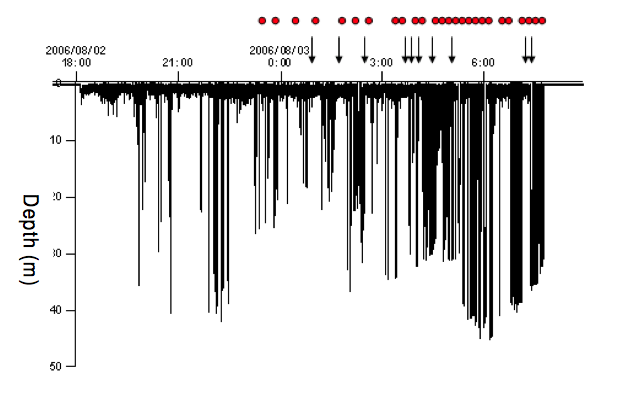
The entire time-depth record during the time the tag was deployed on the dolphin. The tag was on for 13 hrs and 40 minutes. The arrows (  ) mark approximate times of possible foraging events and the red circles ( ) mark times when the dolphin was in acoustic contact with other dolphins, just under nine hours. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
During the 201 days the tag was transmitting our white-beak dolphin spent
44 % of its time within Faxaflói sector, 28 % in the Westfjords sector, 18 % along the south coast and 11 % in the Breiðafjörður sector, and covering an area of roughly 15,000 km2 (Fig. 3). Should this hold in general, it indicates quite a large home range area for Icelandic white-beaked dolphins. Photo identification and observations support the notion that white-beaked dolphins spend much time travelling. Information from photo ID studies of white-beaked dolphins in Faxaflói Bay from 1998 to 2002 indicated an open population with relatively low number of re-sightings in a season. For example in 2002, there were five to nine re-sightings within a 3-month period out of a maximum of 109 different individuals (Rasmussen and Jacobsen, 2003). Bertulli (2010) had a maximum number of 45 inter-annual re- sightings from a catalogue of 245 different individuals from 2007 to 2009 and more than half (60 %) of all re-sighted individuals were seen in the same month. In contrast the Moray Firth bottlenose dolphins in Scotland seem to have a restricted home area since at least 90% of the population (about 130 individuals) visit the bay annually (Wilson et al., 1999). Satellite tagging of Moray Firth dolphins would probably reveal a larger home range. The Sarasota Bay bottlenose dolphins are long-term, year-round residents with a home range of about 125 km2 (Connor et al., 2000).
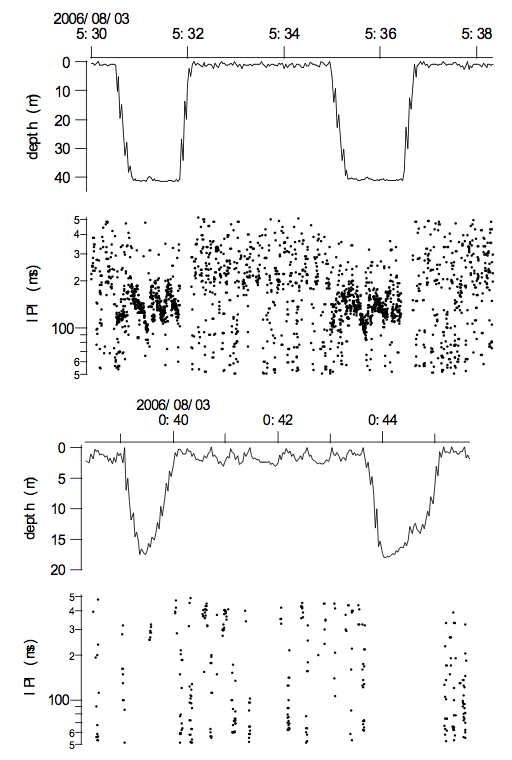
Examples of U-shape dives (upper figure) and V-shape dives (lower figure) including interpulse (click)-intervals (IPI) in ms for both. The jagged traces in the depth curves are caused by the sampling routine of the TDR unit. Note the presumed feeding bouts in U-shaped dives characterized by reducing IPI (see also Figure 7). Note the smaller number of clicks during V-shaped dives comparing with U-shaped dives. Dots near the surface are triggerings caused by splash noise, which were excluded before the analysis in Table 1 using the filtering procedure mentioned in the methods.
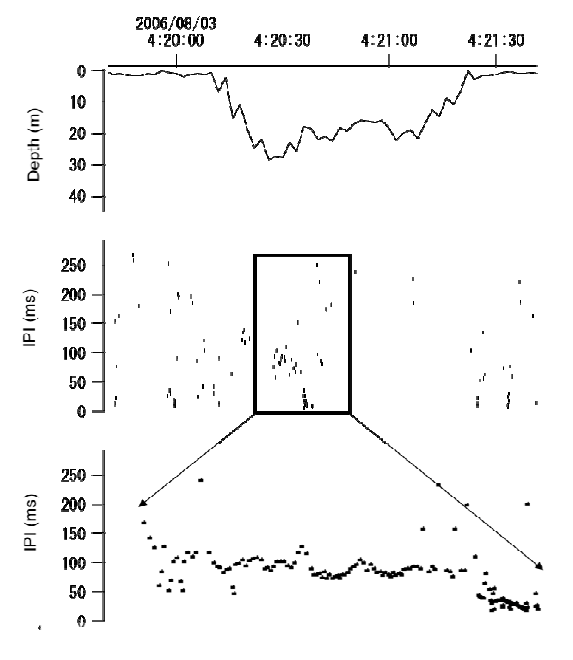
Example of a possible foraging dive. The figure shows an example of a single U- shaped dive, where interpulse (click)-intervals (IPI) are decreasing from 100 ms to 2 ms at a depth of 18 m. The lower part of the figure shows a close up of the decreasing click-intervals during a possible foraging attempt. The jagged traces in the depth curves are caused by the sampling routine of the TDR unit.
White-beaked dolphins are opportunistic feeders and feed on many different species of prey and the movements of our satellite tagged dolphin can in some way be explained by the distribution and hotspots of certain prey species. Visual observations indicate that, among other things, the white-beaked dolphins prey on sandeels, Ammodytes sp., inside Faxaflói Bay during the summer months. As a response to apparent decline in sandeel abundance, the Icelandic Marine Research Institute started a monitoring program of sandeels in 2005 off the south and west coasts of Iceland. The highest concentration of sandeels was found in Faxaflói Bay in 2006 and 2008 with more than 1 fish per m2 (Bogason and Lilliendahl, 2009). This high concentration of sandeels might explain high number of white-beaked dolphin sightings inside Faxaflói Bay during the present project. The peak season for capelin, Mallotus villosus, fisheries along the south coast is in February and March, which correlates with the position of the tagged animal in February so one can speculate that the white-beaked dolphin was feeding on capelin at this time of the year. Previous research has shown that during these months the diet of harbour porpoises consists overwhelmingly of capelin in this area (Víkingsson et al 2003). This is also consistent with Magnúsdóttir (2007) who observed humpback whales (Megaptera novaeangliae) and white-beaked dolphins together with a big flock of gannets (Morus bassanus), kittiwakes (Rissa tridactyla) and fulmars (Fulmarus galcialis) apparently feeding on capelin along the south coast in February 2006. White-beaked dolphins feed also on haddock and cod (Víkingsson and Ólafsdóttir 2004). These are commercially important species exploited year round in all of Iceland including Faxaflói , Breiðafjörður, and the Westfjords.
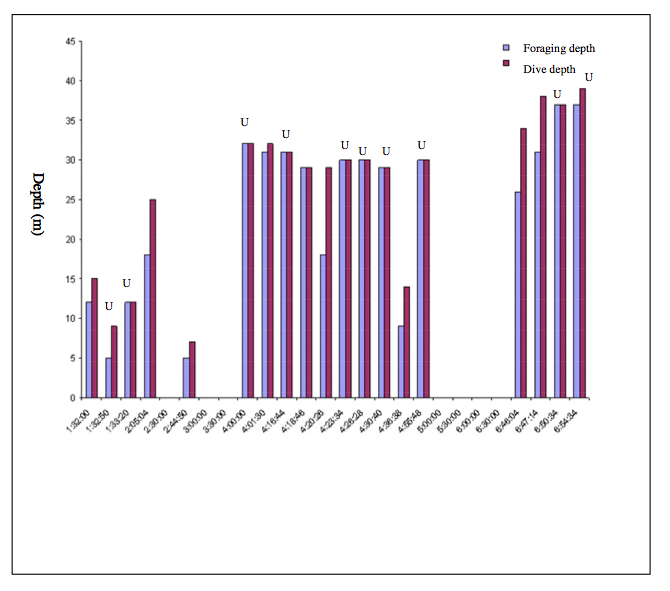
All 19 possible foraging events of the tagged white-beaked dolphin showing the maximum dive and the depth of possible foraging events. Ten of the possible foraging events were in U-shaped dives (marked) and nine in V-shaped dives. Note that about half of the presumed foraging events occurred at the bottom of the dive while the others occur on the descent/ascent. There is a significant difference between U and V- shaped dives when foraging occurs on the descent/ascent (p=0.032), but not at the bottom of the dive.
4.2 Diving and acoustic behaviour of a tagged white-beaked dolphin
This is the first time the echolocation behaviour of a free-ranging dolphin has been studied using a behavioural tag recording diving and acoustics.
The maximum dive depth of our acoustic tagged white-beaked dolphin was 45 m, which is close to the maximum water depth in the inner and coastal part of Faxaflói Bay, about 40 to 50 m. Most likely, the dolphin stayed in this part of the bay and repeatedly dove to the bottom during the time the tag was on (the tag was also retrieved in this part of the bay). Atlantic spotted dolphins (Stenella frontalis) inhabit shallow waters in the Gulf of Mexico and the maximum dive depths were between 40 m and 60 m (Davis et al., 1996). The “bottom time” of our dolphin was between 2 to 78 seconds (mean: 28 s ± 21 s sd). This is a shorter duration than Scott and Chievrs (2009) found for mean bottom time of spotted dolphins (mean: 34 s ± 31 s during day; mean 45 s ± 41 s during night). This difference can be explained by the difference in diving depth. Our dolphin spent 18 % of the time close to the surface (0-2 meter) and 82 % of the time was spent diving (at more than 2 meters). This is comparable to what was found for the white-sided dolphin and the bottlenose dolphin. The white- sided dolphin spent about 89 % of the time submerged (Mate et al., 1994) and the bottlenose dolphin was submerged for 87.1% of the total time (Mate et al., 1995).
Our tagged white-beaked dolphin emitted clicks nearly continuously with no periods of long silence. Clicks from neighbouring dolphins were first recorded about five hours after tagging and for the remaining 8 hours and 40 minutes (Fig. 4). Whistle vocalizations could not be registered by our system.
White-beaked dolphins were using click intervals at the bottom of the dive with a mean interval of 177 ms in V-shaped dives and mean 134 ms in U-shaped dives. In search distance this corresponds to 132 m and 101 m respectively when taking the two way travel time into account. In comparison harbour porpoises use click intervals varying between 30 ms and 150 ms during search phase (Linnenschmidt et al., in press) corresponding to a search distance of 23 m to 113 m. Harbour porpoises emit narrow band clicks with a 3 dB beam width of 22 degrees (Au et al., 1999) at maximum source levels (Villadsgaard et al. 2007) about 20 dB weaker than those of white-beaked dolphins. White-beaked dolphins emit broadband clicks (Rasmussen and Miller, 2002) with a 3dB beam width of 8 degrees (Rasmussen et al., 2004). Using the beam width and search distances of 23 m a porpoise would ensonify a surface of 61 m2 and at a search distance of 113 m this would be 1479 m2. For the white-beaked dolphin to ensonify similar search surfaces it would have to be at search distances of 64 m and 310m. Thus the harbour porpoise can cover much larger search surfaces at shorter distances compared to the white-beaked dolphin, assuming the animals can hear the echoes of prey at these distances. If the argument holds true an overlap of foraging strategies could be avoided by two odontocete species competing for the same prey in the same coastal waters.
Is there a reason for the different dive profiles? Westgate et al. (1995) suggest that U-shaped dives to be used during foraging and V-shaped dives during travelling. Our tagged white-beak dolphin apparently foraged in both U- and V-shaped dives (Fig. 8). It spent more time in U-shaped diving and used clicks at a higher rate and shorter intervals compared to the V-shaped dives (Table 3). All presumptive foraging activity occurred between 1:30 am and 7 am. Many of these foraging events were near the bottom of Faxaflói Bay, about 40 to 50 m. Since there was a population boom of sandeels in 2006 when our study took place one could speculate that our tagged white-beaked dolphin preyed on these. Sandeels in the North Sea burrow into the sediment at night and move up in the water column during the day (Macer et al., 1966). The behaviour of sandeels in Iceland during the summer with limited darkness is not known. In any case the dolphin could capture sandeels in the water column or dig them out of the sediment in a method used by bottlenose dolphins and spotted dolphins described by Rossbach and Herzing (1997) and Herzing et al. (2003). Thus we can conclude that both types of dives are used for presumptive foraging. Perhaps V-shaped dives are also used during travelling since the click intervals are significantly longer than those used in U-shaped dives, making it possible for the animal to probe at longer distances.
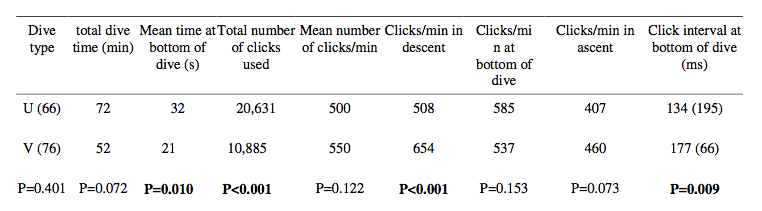
U and V-shaped dives with click statistics for each dive type during the 13 h and 40 m of behavioral recordings from a tagged white-beaked dolphin. Numbers in parentheses are n-values. Bold lettering indicates significant differences. Times spent in the ascent and descent phases as well as the click intervals in these phases were not significantly different in U-shaped and V-shaped dives.
5. Conclusions
This first study of a satellite tagged white-beaked dolphin showed that during the 201 days of tracking the animal moved over 5000 km in coastal waters, but spent nearly half of the time in the bay where it was captured. Our result support earlier anecdotal observations of movements and the year-round presence of white-beaked dolphins in Icelandic waters. The diving behaviour of our tagged animal resembles that of other monitored dolphins. It used echolocation nearly constantly and met up with other odontocetes, presumably white-beaked dolphins, about four hours after release and stayed with these until the tag came off. Our tagged dolphin showed presumed foraging activity in both U-shaped and V-shaped dives, probably near or at the sea bottom. The search range used during U-shaped dives was shorter than that used during V-shaped dives, based upon inter-click intervals. Because of this and the predominance of V-shaped diving, this dive type could be used during travelling.
Acknowledgement
Special thanks to Paul Nachtigall, Aran Mooney, Kristin Taylor, Meike Linnenschmidt, Jeff Foster, Guðlaugur Bjarnasson, Kristinn Guðlaugsson for help during the entire project. Also thanks to Katja Vinding Petersen from Zoovisions for filming the project. Thanks to Signe Sveegaard for creating the map for Figure 3. For financial support we thank the Danish National Research Council, the Danish Research Council for Natural Sciences, the Icelandic Research Fund (RANNIS) and Japan Science and Technology Agency, CREST. We thank constructive comments from two referees that helped improve the manuscript.
References
Akamatsu, T., Wang, D., Wang K. 2005a. Off-axis sonar beam pattern of free-ranging finless porpoises measured by a stereo pulse event data logger. J. Acous. Soc. Am. 117, 3325-3330.
Akamatsu, T., Wang, D., Wand, K., Naito, Y., 2000. A method for individual identification of echolocation signals in free-ranging finless porpoises carrying data loggers. J. Acous. Soc. Am. 108, 1353 – 1356.
Akamatsu, T., Wang, D., Wang, K., Naito, Y., 2005b. Biosonar behaviour of free- ranging porpoises. Proc.R. Soc.London Ser B 272, 797 – 801.
Akamatsu, T., Wang, D., Wang, K., Li, S., Dong, S., 2010. Scanning sonar of rolling porpoises during prey capture dives. J. Exp. Biol. 213, 146-152.
Akamatsu, T., Matsuda, A., Suzuki, S., Wang, D., Wang, K., Suzuki, M., Muramoto, H., Sugiyama, N., Oota, K. 2005c. New Stereo Acoustic Data logger for free-ranging dolphins and porpoises. Mar. Technol. Soc. J. 39, 3-9.
Akamatsu, T., Teilmann, J., Miller, L.A., Tougaard, J., Dietz, R., Wang, D., Wang, K., Siebert, U., Yasuhiko, N. 2007. Comparison of echolocation behaviour between coastal and riverine porpoises. Deep Sea Res. Part II 54, 290-297.
Aguilar, S., N., Johnson, P.M., Madsen, P.T., Díaz, F., Domínguez, I., Brito, A., Tyack, P. 2008. Cheetahs of the deep sea: deep foraging sprints in short-finned pilot whales off Tenerife (Canary Islands)”. J. Anim. Ecol., 77, 936-947.
Au, W.W.L. 1993. The Sonar of the dolphins. Springer-Verlag, New York, Inc.
Au, W.W.A., Kastelein, R.A., Rippe, T., Schoonemann, N.M. 1999. Transmission beam pattern and echolocation signals of a harbour porpoise (phocoena phocoena). J. Acous. Soc. Am. 106, 3699-3705.
Au, W. W. L., Herzing, D. L. 2003. Echolocation signals of wild Atlantic spotted dolphin (Stenella frontalis). J. Acous. Soc. Am. 113, 598-604.
Au, W.W.A., Popper, A.N., Fay, R.R. 2000. Hearing by whales and dolphins.
Springer-Verlag, New York, Inc.
Baird, R.W., Fabrizio Borsani, J. F., Hanson, M, B., Tyack, P.L. 2002. Diving and night-time behavior of long-finned pilot whales in the Ligurian Sea. Mar. Ecol. Prog. Ser. 237, 301-305.
Baird, R.W., Ligon, A.D., Hooker, S.K., Antoinette M. Gorgone, A. M., 2001. Subsurface and nighttime behaviour of pantropical spotted dolphins in Hawai’i. Can. J. Zool. 79, 988-996.
Baird, R.W., Webster, D.L., McSweeney, D.J., Ligon, A.D., Schorr, G.S., Barlow, J., 2006. Diving behaivour of Cuvier’s (Ziphius cavirostris) and Blainville’s (Mesoplodon densirostris) beaked whales in Hawai’i. Can. J. Zool. 84, 1120-1128.
Baird, R.W., Webster, D.L., Schorr, G.S., McSweeney, D.J. Barlow, J. 2008. Diel variation in beaked whale diving behavior. Mar. Mammal Sci. 24, 630-642.
Bertulli, C.G. 2010. Minke whale (Balaenoptera acutorostrata) and white-beaked dolphin (Lagenorhynchus albirostris) feeding behaviour in Faxaflói Bay, south-west Iceland. Master thesis. Institute of biology, University of Iceland, Reykjavik, Iceland.
Bogason, V. Lilliendahl, K., 2009. Rannsóknir á sandsíli / An initiation of sandeel monitoring in Iceland. Hafrannsóknir nr. 145, pp 36-41 (In Icelandic).
Connor, R.C., Wells, R.S., Mann, J. Read, A.J. 2000. The bottlenose dolphin. Social Relationship in a Fission-Fussion Society. In: Mann, J., Connor, R.C., Tyack, P.L. Whitehead, H. (Eds.). Cetacean Societies, field studies of dolphins and whales. The University of Chicago Press, pp. 91-126.
Davis, R.W., Worthy, G.A.J., Würsig, B., Lynn, S.K., Townsend, F.I., 1996. Diving behavior and at-sea movements of an Atlantic spotted dolphin in the Gulf of Mexico. Marine Mammal Science Vol. 12, 569-581.
Deruiter, S.L., Bahr, A., Blanchet, M-A, Hansen, S.H., Kristensen, J.H., Madsen, P.T., Tyack, P.L. and Wahlberg, M., 2009. Acoustic behaviour of echolocating porpoises during prey capture. J. Exp. Biol. 212, 3100-3107.
Dietz, R., Heide-Jørgensen, M.P. 1995. Movements and swimming speed of narwhals, Monodon monoceros, equipped with satellite transmitters in Melville Bay, Northwest Greenland. Can. J. Zool. 73, 2120-2132.
Dietz, R., Heide-Jørgensen, M. P., Richard, P., Acquarone, M. 2001. Summer and fall Movements of Narwhals (Monodon monoceros) from Northeastern Baffin Island towards Northern Davis Strait. Arctic 54, 244-261.
Griffin, D. R. 1958 Listning in the dark: the acoustic orientation of bats and men. New Haven, CT: Yale University Press.
Gunnlaugsson, T., Sigurjónsson J., Donovan G.P., 1988. Aerial survey of Cetaceans in the coastal waters off Iceland, June-July 1986. Rep. Int. Whal. Comm. 38, 489-500.
Gunnlaugsson T., Sigurjónsson J. 1990. NASS-87: Estimation of Whale abundance based on observations made onboard Icelandic and Farose survey vessel. Rep. Int. Whal. Comm. 40, 571-580.
Herzing, D.L., Moewe, K. and Brunnick, B.J. 2003. Interspecies interactions between Atlantic spotted dolphins, Stenella frontalis and bottlenose dolphins, Tursiops truncatus, on Great Bahama Bank, Bahamas. Aquat. Mamm. 29, 352-341.
Hobbs, R.C., Laidre, K.L., Vos, D.J., Mahoney, B.A., Eagleton, M. 2005. Movements and Area Use of Belugas, Delphinapterus leucas, in a Subarctic Alaskan Estuary. Arctic 58, 331–340.
Hooker, S.K., Baird, R.W. 2001. Diving and ranging behaviour of odontocetes: a methodological review and critique. Mammal Rev. 31, 81–105.
Johnson, M., Tyack P.L. 2003. A digital acoustic recording tag for measuring the response of wild marine mammals to sound. IEEE J.Oceanic Eng. 28, 3–12.
Johnson, M., Aguilar de Soto, N. Madsen, P.T. 2009. Studying the behaviour and sensory ecology of marine mammals using acoustic recording tags: a Review. Mar. Ecol. Prog. Ser. 395, 55-73.
Johnson, M, Madsen, P.T., Zimmer, W.M.X., Aguilar de Soto, N., Tyack, P.L. 2006. Foraging Blainville’s beaked whales (Mesoplodon densirostris) produce distinct click types matched to different phases of echolocation. J. Exp. Biol. 209, 5038-5050.
Johnson, M,, Madsen, P.T., Zimmer, W.M.X., De Soto, N.A., Tyack, P.L. 2004. Beaked whales echolocate on prey. Proc. R. Soc. London B, Ser. B 271, 383–386.
Johnson, M, L. Hickmott, LS, Aguilar Soto, N and Madsen, PT 2007. Echolocation behaviour adapted to prey in foraging Blainville’s beaked whale
(Mesoplodon densirostris). Proc. R. Soc. London, Ser. B 275, 133-139.
Linnenschmidt, M., Teilmann, J., Akamatsu, T., Dietz, R., Miller, L.A. (in press). Biosonar, dive and foraging activity of satellite tracked harbor porpoises. Mar. Mamm. Sci. 121 DOI: 10.1111/j.1748-7692.2012.00592.x.
Macer, C.T. 1966. Sandeels (Ammodytidae) in the south-western North Sea; their biology and fishery. Fish. Invest. Ser. II 24, 1-54.
Madsen, P.T., Johnson M., Aguilar Soto N., Zimmer W.M.X, and Tyack P. 2005. Biosonar performance of foraging beaked whales, Mesoplodon densirostris J. Exp. Biol. 208, 181-194.
Magnúsdóttir, E.E. 2007. Year-round distribution and abundance of white-beaked dolphins (Lagenorhynchus albirostris) off the southwest coast of Iceland, Master Paed., Institute of Biology, University of Iceland, Reykjavik, Iceland.
Mate, B.R., Stafford, K.M., Nawojchik, R., Dunn, L.J. 1994. Movements and dive behavior of a satellite-monitored Atlantic white-sided dolphin (Lagenorhynchus acutus) in the gulf of Mar. Mamm. Sci. 10, 116-121.
Mate, B.R., Rossbach, K.A., Nieukirk, S.L., Wells, R.S., Irvine, A.B., Scott, M.D. Read, A.J. 1995. Satellite-monitored movements and dive behavior of a bottlenose- dolphin (Tursiops truncates) in Tampa Bay, Florida. Mar. Mamm. Sci. 11, 452-463.
Miller, L.A. 2010. Prey capture by harbor porpoises (Phocoena phocoena):
A comparison between echolocators in the field and in captivity. J. Mar. Acoust. Soc. 2010. 37, 156-168.
Miller, L.A., Pristed, J., Møhl, B., Surlykke, A. 1995. The click-sounds of narwhals (Monodon monoceros ) in Inglefield Bay, Northwest Greenland. Mar. Mamm. Sci., 1995. 11, 491-502.
Miller, P.J.O., Johnson, M. Tyack, P. 2004. Sperm whale behaviour indicates the use of echolocation click buzzes ‘creaks’ in prey capture. Proc. R. Soc. London Ser. B 271, 2239–2247.
Mooney, T.A., Nachtigall, P., Taylor, K.A., Rasmussen, M.H., Miller, L.A. 2009. Auditory temporal resolution of a wild white-beaked dolphin (Lagenorhynchus albirostris). J. Comp. Physiol. 195, 375-384.
Nachtigall, P.E., Mooney, T.A., Taylor, K.A., Miller, L.A., Rasmussen, M.H., Akamatsu, T., Teilmann, J., Linnenschmidt, M., Vikingsson, G.A. 2008. Shipboard measurements of the hearing of white-beaked dolphins, Lagenorhynchus albirostris. J. Exp. biol. 211, 642-647.
O’Connor, S., Campbell, R., Cortez, H., Knowles, T., 2009. Whale Watching Worldwide: tourism numbers, expenditures and expanding economic benefits, a special report from the International Fund for Animal Welfare, Yarmouth MA, USA, prepared by Economists at Large.
Otani, S., Naito, Y., Kawamura, A, Kawasaki, M, Nishiwaki, S, Kato, A., 1998. Diving behaviour and performance of harbour porpoises, Phocoena phocoena, in Funka Bay, Hokkaido, Japan. Mar. Mamm. Sci. 14, 209-220.
Perrin, W.F., Evans, W.E. and Holts, D.B. 1979. Movements of Pelagic Dolphins (Stenella Spp.) in the Eastern Tropical Pacific as Indicated by Results of Tagging, With Summary of Tagging Operations, 1969-76. Report. NMFS SSRF-737, 1-14.
Pike, D.G., Paxton, C.G.M., Gunnlaugsson, T. Vikingsson, G.A. 2009. Trends in the distribution and abundance of cetaceans from aerial surveys in Icelandic coastal waters, 1986-2001. North Atlantic Marine Mammal Commission Scientific Publications Vol. 7, pp 117 – 142.
Rasmussen, M.H. 1999. Sound production, behaviour and distribution of white- beaked dolphins (Lagenorhynchus albirostris). Master thesis (in Danish). University of Southern Denmark, Odense University.
Rasmussen, M.H. 2004. A study of communication and echolocation sounds recorded from free-ranging white-beaked dolphins (Lagenorhynchus albirostris) in Icelandic waters. Ph.D. thesis, University of Southern Denmark, Odense University.
Rasmussen, M.H, Miller, L.A. 2002. Whistles and clicks from white-beaked dolphins, Lagenorhynchus albirostris recorded in Faxaflói Bay. Aquat. Mamm. 28, 78-89.
Rasmussen, M.H., Jacobsen, T. 2003. Photo-identification of White-beaked dolphins in Icelandic waters. In: ECS proceedings 2003, Las Palmas, Gran Canaria.
Rasmussen, M.H., Miller, L.A. Au, W.W.L. 2002. Source levels of clicks from free- ranging white beaked dolphins Lagenorhynchus albirostris Gray 1846) recorded in Icelandic waters. J. Acoust. Soc. Am. 111, 1122-1125.
Rasmussen, M.H., Wahlberg, M., Miller, L.A. 2004. Estimated transmission beam pattern of clicks recorded from free-ranging white-beaked dolphins (Lagenorhynchus albirostris). J. Acoust. Soc. Am. 116, 1826-1831.
Rasmussen MH, Lammers M, Beedholm K, and Miller LA 2006. Source levels and harmonic content of whistles in white-beaked dolphins (Lagenorhynchus albirostris). J. Acoust. Soc. Am. 120: 510-517.
Read, A.J., Westgate, A.J. 1997. Monitoring the movements of harbour porpoises (Phocoena phocoena) with satellite telemetry. Mar. Biol. 130, 315-322.
Reeves, R.R., Smeenk, C., Kinze, C.C., Brownell, R.L., Lien, Jr., Lien, J. 1999. White-beaked dolphin, Lagenorhynchus albirostris Gray, 1846. In: Ridgeway, S.H., Harrison, S.R. (Eds.). Handbook of Marine Mammals Vol. 6. Academic Press, San Diego, pp 1-30.
Rossbach, K. A., Herzing, D.L. 1997. Underwater observations of benthic-feeding bottlenose dolphins (Tursiops truncatus) near Grand Bahama Island, Bahamas. Mar. Mamm. Sci. 13, 498 – 504.
Salo, K. 2004. Distribution of cetaceans in Icelandic waters. MSc thesis, Institute of Biology, University of Southern Denmark, Odense, Denmark.
Scott, M.D., Chivers, S.J. 2009. Movements and diving behavior of pelagic spotted dolphins. Mar. Mamm. Sci. 25, 137–160.
Scott, M.D., Wells, R.S., Irvine, A.B., Mate, B.R. 1990. Tagging and Marking Studies on small cetaceans. In: Leatherwood, S., Reeves, R.R. (Eds.). The Bottlenose dolphin. Academic Press. pp 489-514.
Shapiro, A. 2006. Preliminary evidence for signature vocalizations among free- ranging narwhals (Monodon monoceros). J. Acous. Soc. Am. 120, 1695-1705.
Sigurjónsson, J. Vikingsson, V. 1997. Seasonal abundance and estimated food consumption by cetaceans in Icelandic and adjacent waters. J. Northwest Atl. Fish. Sci. 22, 271-287.
Svegaard, S., Teilmann, J., Berggren, P., Mouritsen, K.N., Gillespie, D.,Tougaard, J. 2011. Acoustic surveys confirm the high-density areas of harbour porpoises found by satellite tracking. ICES J. Mar. Sci. 68, 929, 936.
26
Suydam, R.S., Lowry, L.F., Frost, K.J., O’Corry-Crowe, G.M. Pikok, D. 2001. Satellite Tracking of Eastern Chukchi Sea Beluga Whales into the Arctic Ocean. Arctic 54, 237–243.
Sæmundsson, B. 1939. Mammalia. The Zoology of Iceland. Volume IV (76).
Tetley, M.J., Wald, E., Björgvinsson, Á., 2006. White-Beaked Dolphin Movements around Iceland: Evidence from Photo-Identification Studies. Poster Presentation, American Cetacean Society Conference, Ventura, 2006
Verfuss, U., Miller, L.A, Pilz, P.K.D. Schnitzler, H.-U. 2009. Echolocation by two foraging harbour porpoises (Phocoena phocoena). J. Exp. Biol. 212, 823-834.
Villadsgaard, A., Wahlberg, M., Tougaard, J., 2007. Echolocation signals of wild harbour porpoises, Phocoena phocoena. J. Exp. Biol. 210, 56-64.
Víkingsson, G.A., Ólafsdóttir, D. and Sigurjónsson, J. 2003. Geographical, and seasonal variation in the diet of harbour porpoises (Phocoena phocoena) in Icelandic coastal waters. North Atlantic Marine Mammal Comission Scientific Publications vol. 5, 243-270.
Víkingsson, G.A. Ólafsdóttir, D. 2004. Hnýðingur (White-beaked dolphin). In: Hersteinsson, P. (Ed). Íslensk Spendýr (Icelandic Mammals). Vaka-Helgafell, Reykjavík, Iceland, pp 154 – 157.
Wahlberg, M, Jensen, FH, Natacha Aguilar Soto, NA, Beedholm, K, Bejder, L, Oliveira, C, Rasmussen, M, Simon, M, Villadsgaard, A and Madsen, P 2011. Source parameters of echolocation clicks from wild bottlenose dolphins (Tursiops aduncus and Tursiops truncatus). J. Acoust. Soc. Am. 130, 2263-2274.
Westgate, A.J., Read, A.J., Berggren, P., Koopman, H.N., Gaskin, D.E. 1995. Diving behaviour of harbour porpoises, Phocoena phocoena” Can. J. Fish. Aquat. Sci. 52, 1064-1073.
Wilson, B, Hammond, P.S., Thompson, P.M. 1999. Estimating size and assessing trends in a coastal bottlenose dolphin population. Ecol. Appl. 9, 288-300.
Winslade, P. 1974. Behavioral studies on the lesser sandeel Ammodytes marius (Raitt). The effect of food availability on activity and the role of olfaction in food dectection. J. Fish. Bull. 6, 565-576.
Zimmer, W.M.X., Johnson, M.P., Madsen, P.T., Tyack, P. 2005. Echolocation clicks of free-ranging Cuvier’s beaked whales (Ziphius cavirostris). J. Acous. Soc. Am. 117, 3919 – 3927.
